On Microbiome Prescription we have Medical Conditions with Microbiome Shifts from US National Library of Medicine with some 91 entries. It would be good for everyone if we increase the number of entries.
YOU can help make it happen!
- Find a condition of concern to you or someone you know.
- Go to https://pubmed.ncbi.nlm.nih.gov/
- Type in the name with ” 16s microbiome “, for example, with “pregnancy”

Click Search. you will hopefully get a few dozen (or hundreds) of studies.
Next is the long haul part. Go thru them (Example URL).
- For many, the title alone tells you to skip it!! For example Interaction between Microbes and Host in Sow Vaginas in Early Pregnancy.
- Remember, we are not dealing with vaginal microbiome on the site, so you can skip all of those too!
- Others may be interesting: Effects of galactooligosaccharides on maternal gut microbiota, glucose metabolism, lipid metabolism and inflammation in pregnancy: A randomized controlled pilot study.
- Look at the summary or the full study if available
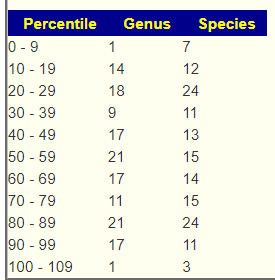
- Look for bacteria names. We need to know the name and how they shift. i.e.
And compared with the placebo group, the GOS group had a higher abundance of Paraprevotella and Dorea, but lower abundance of LachnospiraceaeUCG_001.
- Then comes the thinking part. Trying to describe the results. In this case, we see that the focus is “gestational diabetes mellitus (GDM)”
- If not a good fit for your focus, add it to a new list to come back to later.
In the above case, a new search (gestational diabetes mellitus 16s microbiome) returned 70+ studies. This is a good candidate for a new collection to add.
Sending the Information to Me
I would suggest putting the links (i.e. like https://pubmed.ncbi.nlm.nih.gov/36471554/ ) in a text file or excel and sending to me.
The information sent (when added to database) is available to everyone! This is citizen science! As I remarked to someone earlier today “I do not have a business model, I have a pro bono model“.


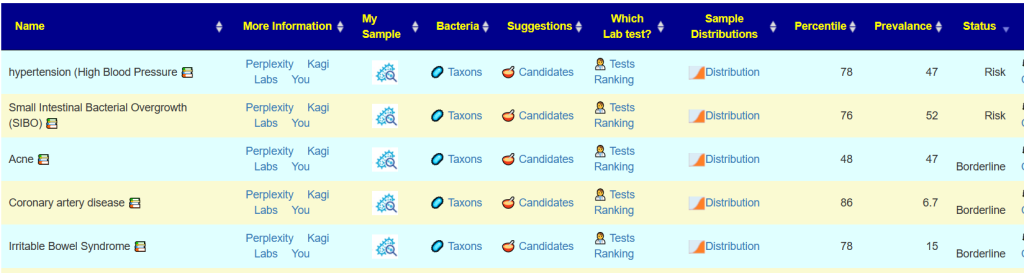
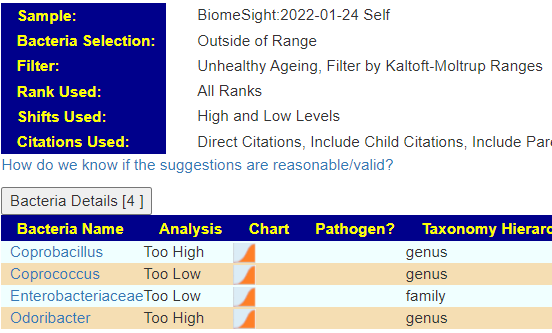
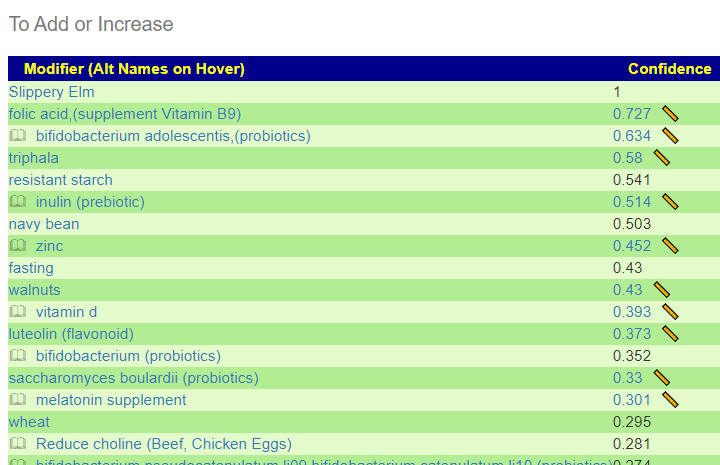
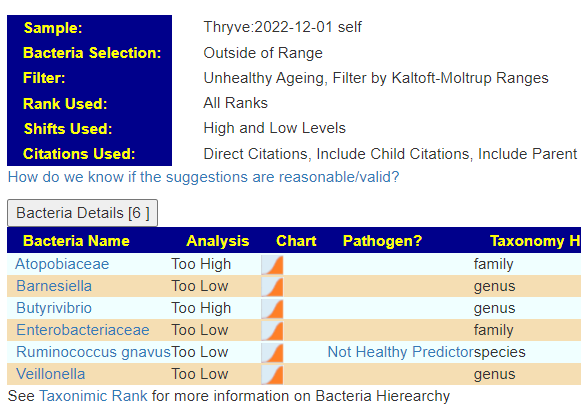
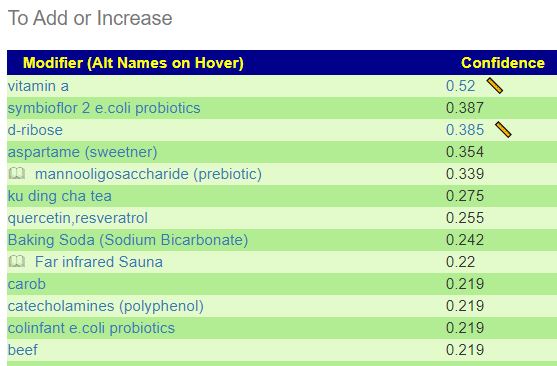
The encoded data can be used to evaluate yours and others microbiome against the patterns reported.

That is it!!! You spend the time so others do not have to and can act on their challenges with better information!
Email the lists to Research@microbiomeprescription.com






Recent Comments